Social calls of the Nathusius' pipistrelle (Pipistrellus nathusii)
Read more about how I categorized the unusual social calls of the Nathusius's pipistrelle in this blog post.
A collection of the part A0, D0 and E motifs of the type D social call of the Nathusius' pipistrelle can be found on this page.
The following recording contains type A, B, C and D social calls. Hence why it is a great example to show as a start on this page. It is presumed that the bats emitted these calls stationairy from within the roost - possibly a mating roost.


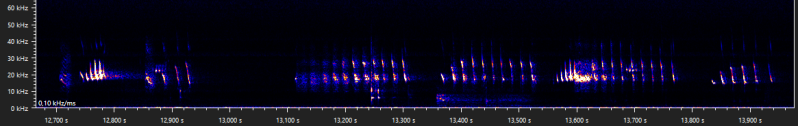


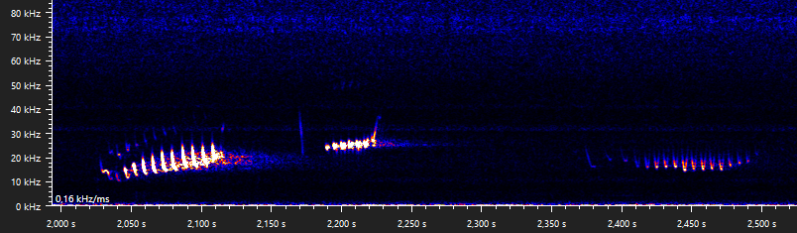




In the last few snippits of the previous examples, the typical part A of the type D social call appears to merge into a more nasal sound. Another bat doing something similar has been recorded before:






Date of recording: 2024-08-24 Recorded by: Niels Jansen Audio file: 932560
The following type B and type D social calls were recorded from a stationary Nathusius' pipistrelle inside a bat box on the gable of a house.



And these type B social calls (between 6,000s and 6,400s) were present within a syntax of otherwise type D social calls of an advertising male:

Date of recording: 2024-08-28 Recorded by: Kaia Pieters Audio file: 936333
More similar type B were recorded (alongside part D motifs):

Date of recording: 2024-08-22 Recorded by: Molito Fego Audio file: 931602
And these:

Date of recording: 2024-09-16 Recorded by: Isis Knijff Audio file: 935111
And these type B and D social calls were emitted by a stationary advertising male:

Date of recording: 2024-09-17 Recorded by: Levi Laluan Audio file: 935731
More type B and D social calls emitted by a stationary advertising male:
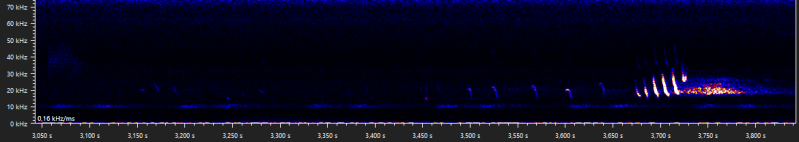
Date of recording: 2024-08-27 Recorded by: Cas van Joolingen Audio file: 936337
And perhaps these social calls, also emitted between type D social calls of an advertising male, could be type B as well. Even though some calls seem to lean towards part D of the type D social calls. However, these calls have a distintive alternating "tick-tock" sound to them.


Here is another example of what I would call the "tick-tock" sound (between 24,530s and 24,860s):

Date of recording: 2024-09-18 Recorded by: Aielynn Kleian Audio file: 1040338
And this one might be as well (between 6,380 and 6,540s):

Date of recording: 2024-09-16 Recorded by: Levi Laluan Audio file: 955941
The next two examples are the most general variant of the most common type D social call of the Nathusius' pipistrelle. It exists out of three parts, that we will call part A (multi-component), part B (single FM component) and part C (fast trill). Part A can differ in the number of components. Part B and part C are variable in appearance as well and can differ between individuals. There Nathusius' pipistrelle can also shuffle the different parts to create new songs. The sequence and different combinations of parts can convey a more complex message to other bats (Middleton et al., 2020).
Five-component variant of Part A:

Date of recording: 2024-05-18 Recorded by: Niels Jansen Audio file: 911324
Seven-component variant of part A:

Date of recording: 2024-05-26 Recorded by: Niels Jansen Audio file: 911808
Part A can also contain extra components that go up to 70 - 80 kHz in frequency at the end of part A's regular sequence. In existing literature, the main motif of part A is called A1 and the extra components at the end of the motif are called A2 (Hargreaves et al., 2017).

Date of recording: 2024-06-16 Recorded by: Sander Boersma Audio file: 913412
A part A1 can also contain a pre-part that I will call part A0. These are some extra components in front of the main motif (see the two snippits on the right). In the example below, I suspect that this part A0 is used as a stand-alone motif (or simply cut off after) inbetween the other parts.



Date of recording: 2024-06-26 Recorded by: Menno van Berkum Audio file: 919195
A Nathusius' pipistrelle emitting part A motifs of the type D social call. If this is the only motif clearly visible, confusion with the type D social call of the common pipistrelle may occur on first sight. However, the Nathusius' pipistrelle tends to go up in frequency with each components, creating a shape similar to "◢".

Date of recording: 2024-06-26 Recorded by: Cas van Joolingen Audio file: 917084
This example contains a "shuffled" sequence of parts. These three syntaxes each consist out of the sequence part B-C-B.

Date of recording: 2024-06-14 Recorded by: Olivier van Aken Audio file: 913613
The sequence part B-C:

The sequence part A-B:

Date of recording: 2024-05-27 Recorded by: Hubert Matuszewski Audio file: 911552
In this example the bat emitted the motif part B zeven times in a row. Without the presence of other type D motifs, the part B motif is a type C social call on itself.

And in the same recording, the following social call sequence is also present: part A, part B, part C, part B and a new kind of motif that exists out of multiple FM components. This is a motif that I will call part E.

Date of recording: 2024-05-20 Recorded by: Gijs Peters Audio file: 911550
An example of a Nathusius' pipistrelle emitting the following sequence of motifs: part B, part B and C, part B and E.

Date of recording: 2024-06-14 Recorded by: Nils Reinerie Audio file: 913335
The following snippit contains the motifs: part A+B+E, part B+C and part B+C.

Date of recording: 2024-05-28 Recorded by: Anice Hut Audio file: 911811
These social calls of the Nathusius' pipistrelle were recorded near a foraging area. It is most likely agonistic behaviour of the Nathusius' pipistrelle towards the common pipistrelle in "his" foraging area. Among other calls, the recording contains the following sequence of motifs: Part A, B - B, E - B, C.

Date of recording: 2024-07-02 Recorded by: Gijs Peters Audio file: 918333
Then you might wonder about the motif part D. While part E is made up out of multiple FM components, part D is made out of multiple downward FM sweeps with a relatively longer qCF ending.
The following two recordings are calls from a Nathusius' pipistrelle that was emitting these calls stationery from behind a gutter. The call between 5,900 s and 6,960 s (the first snippit on the right below) is visiually and audibly similar to the faint (or perhaps 'non-joined') slow trill, followed by three FM-qcF components, recorded by Lisa on 2024-06-14 (recording 913306), and is thus suspected to be the same type of call as that one - not a part D.









The following part D motif were recorded by Niels at the same location, the same evening:



A very beautiful example of a part D call:

Date of recording: 2024-08-19 Recorded by: Levi Laluan Audio file: 930316
All previous social calls recorded by Niels were recorded near a roost of Nathusius' pipistrelle where at least ten bats were seen entering the roost. One more oddly shaped call was recorded at this location. It is a call in the shape of a walking stick, present at 23,550 s in the following snippit of the recording. Modified echolocation calls with hooks emitted by Nathusius' pipistrelle were also documented by Jahelková (2011).

Date of recording: 2024-05-26 Recorded by: Niels Jansen Audio file: 912056
More modified echolocation calls were recorded during mating season, together with some unusual social calls. Note the five hooked echolocation calls between 4,600s and 5,200s in the first snippit and one more between 2,750s and 2,800s in the second snippit:


Hooked echolocation calls that are more similar to the walking stick shaped social calls (similar to those of the pond bat and Daubenton's bat) are present in the following snippits. Sometimes it appears to be the arch only or even just the ascending part of the arch.









In my opinion, the modified echolocation calls of the previous recordings also explain the shape of the following calls. Two bats were seen flying through the street. One of the bats tapped a bat box on the wall on its way. The recording starts with type D social calls of the Natusius' pipistrelle. Later in the recording, the echolocation seems to be modified. Even a qCF-FM call is present between 60 and 70 kHz around 19,600s in the recording.


Date of recording: 2024-05-25 Recorded by: Olivier van Aken Audio file: 911361
Our fieldworker Tessa encountered these social calls that look very similar to the part D motifs of the previous recordings by Niels in the vicinity of a Nathusius' pipistrelle roost. However, Tessa saw no behaviour from a bat that would indicate a roost, such as bats entering the building or tapping a possible roost entrance. It is a possibility that the roost's entrance was on the side of the building that was just out of sight.


Some sounds can't be classified to one of the previous described motifs. The recordings of Anice and Lisa are made at the same location, but with almost a month in between.
The first call of the recording of Anice (the snippits on the right) shows a trill. The second snippit shows the motifs part B-E.
The first call shown in the recording of Lisa (the snippits below) is another trill followed by four components that go up in frequency with each component, very similar to a part D motif. Between 8,050 s and 8,110 s (second snippit), there is a faint, or perhaps 'non-joined', trill visible. The third snippit showns another part D motif.


Date of recording: 2024-05-28 Recorded by: Anice Hut Audio file: 911811



Date of recording: 2024-06-14 Recorded by: Lisa Vermaning Audio file: 913306
What is noteworthy, is that another type of call was recorded at the exact same location as the previous two recordings in August (during mating season): a wavy-line call (between 1,850s and 2,000s in the snippit below).

Date of recording: 2024-08-17 Recorded by: Gert-Jan Hendriks Audio file: 928168
The wavy-line call is a warbled qCF call. I recorded these calls the first time myself in May of 2021. The sequence of motifs in the following recording consists out of this wavy-line followed by a part B, a wavy-line followed by a small 'cheep' around 15 kHz and then a part B, a part B-C and a stand-alone part A motif.




Date of recording: 2021-05-10 Recorded by: Sarah Mahie Audio file: 911931 (slightly edited the noise out in the snippits for clarity)



Date of recording: 2024-06-25 Recorded by: Manno Kolvoort Audio file: 916043
Another great example of a wavy-line call (between 1,240s and 1,330s), also used as a motif between other type D social call motifs::

Date of recording: 2024-09-20 Audio file: 1001463
I was eagerly waiting for other people to record these wavy-line calls, since we were collecting recordings on such a big scale this year, and I was not dissapointed. Two wavy-line calls after each other (one starting around 17,650 s and the other around 17,900 s in the snippit below), supplemented by a part B+C+B were recording during spring (outside of mating-season) in the beginning of July.

In the same recording, part E was also used in different call sequences:


Date of recording: 2024-07-02 Recorded by: Chris Johan Diepenmaat Audio file: 918721
This recording was made near an area where a common pipistrelle and a Nathusius' pipistrelle were foraging, as seen from the feeding buzz in the beginning of the recording (not shown). The Nathusius' pipistrelle emits multiple part B motifs. It is suspected that these FM calls are agonistic in nature, since the majority of the recordings where this motif is used as stand-alone type C social calls, is when a foraging Nathusius' pipistrelle is in the vicinity of other species of (foraging) bats. In this particular recording, an interesting type D part D call is present after one of the FM calls.

Date of recording: 2024-06-26 Recorded by: Sanna Gerdes Audio file: 916811
The following recording contains part E and part D motifs. The bat was seen tapping the wall and flew into and later out of it's roost. Continuously flying up and down.

Date of recording: 2024-08-21 Recorded by: Wendy Baars Audio file: 936228
These part D motifs were also emitted while in flight. About five minutes after these two recordings, a serotine bat flew by emitting multiple agonistic calls (914886).








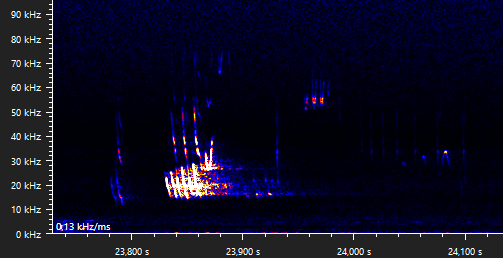
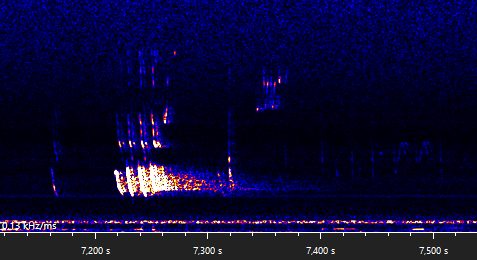
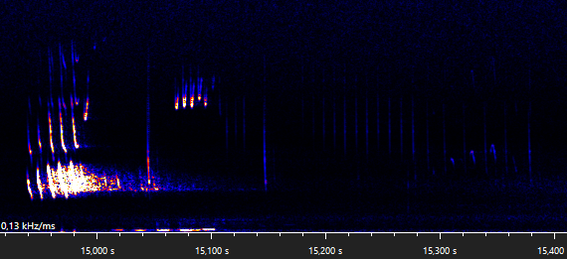
The following recording is made in the same village as the previous recordings, near a foraging area and next to a row of trees that might function as a flying route. The motifs part A, B, C, D, E can be recognized, as well as trills. There seems to be a few extra notes that are occasionally emitted before the main part A(1), a pre-part that I have decided to call part A0.
The calls between 14,750 s and 15,400 s (first snippit below) are hard to distinguish, since at least two bats are emitting calls at the same time. A single bat could not produce a part B and a part A(0) motif at the same time (14,790 s). Since the part A1 motifs are fairly different, I suspect these belong to different individuals. There are four part B motifs present at two different frequencies, two around 19, 5 kHz (bat A) and two around 33,5 kHz (bat B). Since bat A emits a part B motif during the first part A(0) motif, the part A must be emitted by bat B. Since the second part B motif of bat B overlaps with the only present part C motif, we can conclude that the part C motif is emitted by bat A. This leaves us with the following two sequences that, I believe, bat B emitted: part A(0,A)-B and a joined-up part A-B". The second snippet shows another joined-up part A.










Part E, part D and a wavy-line calls were present during this encounter with two Nathusius' pipisrelles.






And the part C motif is going down in frequency in this recording, made a few days earlier at the same location as the previous two recordings:

Date of recording: 2023-08-29 Recorded by: djsaaltink Audio file: 826467
A low-frequency trill that I currently call a "mordent". I consider it a possibility that a mordent is the just the part A0 motif. It was recorded in an area where common pipistrelle, Nathusius' pipistrelle and serotine bat were foraging.

Date of recording: 2024-06-28 Recorded by: Hubert Matuszewski Audio file: 917429
Another mordent that looks very much like the one in the previous recording (between 2,550 s and 2,600 s in the first snippit, between 8,800s and 8,850 s in the second snippit). The same calls were recorded on two locations about 90 meters apart. Both snippits are the same calls from the two different locations. There is one more mordent present (between 3,170 s and 3,250 s in the first snippit, between 9,420 s and 9,500 s in the second snippit). These were recorded in the same area where the trills of 911811 on 2024-05-28 and 913306 on 2024-06-14 were recorded.


The following snippet shows a mordent, a joined-up part A motif and a part B followed by a part D motif.

In the same recording, there is some more proof that some trills originate from 'joined-up' part A motifs as theorized:


Altough, still not all of the trills can be explained that way - perhaps some are 'true trills' or joined-up part E motifs:


I would consider the call between 23,700s and 23,850s to be a wavy-line call:

Date of recording: 2024-08-19 Recorded by: Declan Frerichs Audio files: 929731
A Nathusius' pipistrelle was emitting a series of social calls and tapping the entrance of a bat box on the wall of a buiding before entering the bat box. The type D social calls in the following recording are mainly part A only. Between 1,300 s and 1,900 s and between 3,400s and 4,100 s in te recording there are also some unusual tunes present.
In the recording directly after this, the social calls consisted mainly out of part A and B. In the recording after that one, the social calls consisted out of of part A, B and C.



Date of recording: 2024-06-28 Recorded by: Kaz Veldtrom Audio file: 917393 Video footage by: Kaz Veldtrom
Sometimes, social calls can be very misleading. The following calls took me quite some time to identify. The recording 912750 (not shown) was made around 15 minutes later than the following two recordings and cracked the case for me; a Nathusius' pipistrelle was emitting part B as type C social calls in that recording. I was a little thrown off by the shape, but the type C social calls in a recording of C. Nason (Middleton et al., 2020) would suggest that type C social calls of the Nathusius' pipistrelle can take on this form aswell.


For about 10 minutes, a Nathusius' pipistrelle and common pipistrelle were seen foraging. Feeding buzzes of both species were witnessed. Type C and type D social calls, including a part D motif, are emitted by a Nathusius' pipistrelle. I do not exclude the possibility that some of the type C social calls are from common pipistrelle.




Almost continuously the use of part D motifs:





Sometimes an extra note seems to be present in front of the part A motif:


Date of recording: 2024-08-28 Recorded by: Timo Boer Audio file: 931844
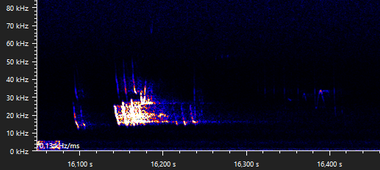





The same bat emitting calls in flight instead of stationary:


Type C social calls, high-frequency as well as low-frequency:

Date of recording: 2024-09-14 Recorded by: Hubert Matuszewski Audio file: 947652
More low-frequency type C social calls emitted by a flying individual:

Date of recording: 2024-08-23 Recorded by: Iris van der Horn Audio file: 931764


Two Nathusius' pipistrelles are present in the following snippit. One of the two bats emits a series of high-frequency type C social calls.

Date of recording: 2024-06-14 Recorded by: Nils Reinerie Audio file: 913335
Two Nathusius' pipistrelles were seen flying close to each other. The echolocation of both bats can be heard up until around 21,300 s in the following recording. Suddenly, it looked like one of the bats (or maybe both attached on each other?) fell down.
There could be two faint calls present at 21,830s and 22,175 s, but they are to faint to hear. Between 22,200 s and 22,450 s there is noise. Directly after at around 22,560s there is a more clear bat call (FM) present with it's peak at 19 kHz. The next call (FM) is at 22,780s, with it's peak around 36 kHz. From that moment on, from 22,830s on forward, it looks like the bat is trying to stabilize it's calls to more regular echolocation calls with the first 'normal-looking' call at 23,095 s around 42,5 kHz. After 23,970s, the echolocation calls of the second bat also become visible again.

Date of recording: 2024-06-23 Recorded by: Thomas Kooiman, Sarah Mahie Audio file: 915774
All recordings are licensed under the following Creative Commons Attribution-NonCommercial-NoDerivs 4.0 license and in courtesy of Sarah Mahie.
All the original video footage can be found on our Youtube channel: United by Ecology
All sonograms are screenshots of the recordings imported in the ultrasound analysis software BatExplorer 2.2 (Elekon, Switzerland).
Literature list:
- Daniel Hargreaves, Helena Jahelkova, Oliver Lindecke and Guido Reiter (2017). Bat Species of the Year 2015: Nathusius’ pipistrelle (Pipistrellus nathusii). Facts compiled for BatLife Europe.
- Helena Jahelková (2011). Unusual social calls of Nathusius' pipistrelle (Vespertilionidae, Chiroptera) recorded outside the mating season. Institute of Vertebrate Biology, Czech Academy of Sciences. Folia Zoologica, 60(1): 25-30. https://doi.org/10.25225/fozo.v60.i1.a4.2011
- Jon Russ and Paul Racey (2007). Species-specificity and individual variation in the song of male Nathusius’ pipistrelles (Pipistrellus nathusii). Behavioral Ecology and Sociobiology, 61(5): 669-677. DOI:10.1007/s00265-006-0295-9
- Jon Russ (2021). Bat Calls of Britain and Europe: a Guide to Species Identification. Pelagic Publishing.
- Niel Middleton (2020). Is That a Bat? A guide to non-bat sounds encountered during bat surveys. Pelagic Publishing.
- Neil Middleton, Andrew Froud and Keith French (2022). Social Calls of the Bats of Britain and Ireland (second edition). Pelagic Publishing.
- Stuart Newson, Neil Middleton and Huma Pearce (2020). The acoustic identification of small terrestrial mammals in Britain. British Wildlife. 32(3).
Create Your Own Website With JouwWeb